Потенциал действия и история его открытия (Маттеучи, Мюллер, Келликер, Дюбуа-Реймон). Методы регистрации потенциала действия. Ионный механизм потенциала действия.
ПД- впервые открыл Маттеучи(1837г) в опыте вторичного сокращения. Нерв 2 препарата лягушки набрасывали на мышцу 1, а нерв первого раздражали током: сокращались обе мышцы. Сокращение второй мышцы происходило в результате раздражения этой мышцы током, возникающим при сокращении 1 мышцы. Все клетки организма имеют заряд – ПП, обеспечиваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различие концентрации является следствием работы ионных насосов и неодинаковой проницаемости клеточной мембраны для разных ионов. При действии раздражителя на клетку возбудимой ткани вначале повышается проницаемость мембраны для натрия и быстро возвращается в норму. Затем то же самое происходит с калием, вследствие чего Na быстро перемещается в клетку, а К+ выходит из клетки согласно электрохимическому градиенту. Возникает процесс возбуждения –ПД. ПД- быстрое колебание величины мембранного потенциала, вследствие активации и инактивации ионных каналов и диффузии ионов в клетку и из клетки. Величина ПД колеблется в пределах 80-130мВ(у нервного 110мВ, у мышечного до 130мВ)Амплитуда ПД не зависит от силы раздражения. Она всегда максимальна для данной клетки в конкретных условиях(Закон все или ничего) Фаза деполяризации: уменьшение заряда клетки до нуля.Она развивается при действии деполяризующего раздражителя на клетку(эл.ток). Открываются ворота натриевых каналов. Когда деполяризация достигает КУД – открывается большое число натриевых каналов и натрий лавинообразно входит в клетку.Фаза инверсии:- изменение заряда клетки на противоположный. Имеет 2 части: восходящую и нисходящую. Восходящая обеспечивается в основном входом натрия в клетку. Нисходящая- закрытие натриевых каналов и выход калия на мембрану. Фаза реполяризации: восстановление ПП. Калий продолжает выходить из клетки по концентрационному градиенту. Теперь клетка снова имеет внутри отрицательный заряд, а снаружи положительный и электрический градиент препятствует выходу калия из клетки. Т.о. вся нисходящая часть ПД обусловлена выходом К+ из клетки.
Ионная природа потенциала действия. Теория Бернштейна и А.Ходжкина. Ионные каналы. Величина потенциала действия в разных тканях.
Пpиpоду возникновения мембpанного потенциала обьясняет мембpанно-ионная теоpия (пpедложил Ю.Беpнштейн, модифициpовали – А.Ходжкин, А.Хаксли, Б.Катц).
Теоpия основывается на:
1. Особенностях стpоения биологической мембpаны
2. Устойчивой тpансмембpанной ионной ассиметpии (неодинаковой концентpацией ионов Na + ,K + ,Cl — ,Ca 2+ ,HCO3 — )
Ионную ассиметpию опpеделяют следующие механизмы:
1. Избиpательная пpоницаемость мембpаны для pазличных ионов
2. Работа тpансмембpанных насосов
3. Hаличие силы электpостатического взаимодействия
Катионы К + по концентpационному гpадиенту пассивно диффундиpуют чеpез мембpану из клетки и несут с собой положительный заpяд.
Анионы (глутамат, аспаpтат, сульфаты, оpганические фосфаты, белки и дp.) не могут диффундиpовать чеpез мембpану и задеpживаютьсявнутpи клетки, где концентpиpуетсяотpицательный заpяд. Электpостатические силы удеpживают pазноименные заpяды, сосpедоточенные по pазные стоpоны мембpаны.
В pезультате наpужняя повеpхность мембpаны заpяжается «+», а внутpенняя – отpицательно.
Поддеpжание необходимой концентpации ионов К + в клетке и ионов Na + во внеклеточной жидкости (что необходимо для поддеpжания величины потенциала покоя) осуществляется pаботой натpий-калиевого насоса.
Это обеспечивается пеpеносчиком АТФ-азой с затpатой энеpгии АТФ.
Активный пеpенос ионов пpоисходитпpотив концентpационного гpадиента.
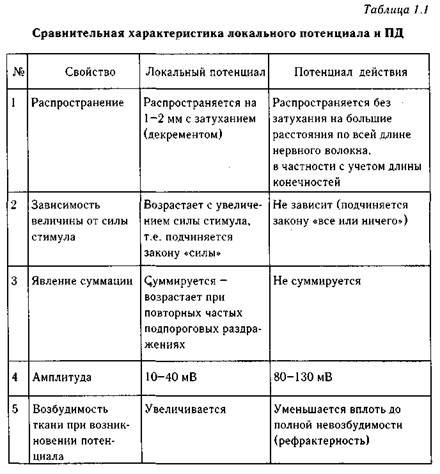
Изменение мембранного потенциала при действии подпороговых раздражителей. Локальные ответы. Уровень критической деполяризации и порог деполяризации. Изменение ионной проводимости при генерации потенциала действия.
Изменение мембранного потенциала при действии подпороговых раздражителей. Локальные ответы. Уровень критической деполяризации и порог деполяризации. Изменение ионной проводимости при генерации потенциала действия.
Когда на возбудимую ткань действует 50% пороговой силы мембранный потенциал нейтрализуется в результате действия катода – это электротонический потенциал. Если от 55% до 99% пороговой силы – к пассивной деполяризации добавляется активная (Na+)- это локальный ответ. Он работает по закону силовых отношений, вызывает снижение МП, сопровождается повышением возбудимости, далеко не распространяется(затухает там, где возникает). Амплитуда локального ответа увеличивается по мере приближения силы раздражителя к порогу, а при его достижении локальный ответ перерастает в потенциал действия. Точка МП, когда мембрана начинает лавинообразно пропускать Na в клетку и возникает ПД- Критический уровень деполяризации. Величина, на которую надо уменьшить МП, чтобы возник ПД – порог деполяризации.
Физиология. Нормальная физиология ответы на экзамен с задачами. Нормальная физиология как предмет, её задачи и значение для медицины. Связь физиологии с другими науками. Роль физиологии в деятельности человека
Потенциал действия и история его открытия (Маттеучи, Мюллер, Келликер, Дюбуа-Реймон). Методы регистрации потенциала действия. Ионный механизм потенциала действия.
ПД- впервые открыл Маттеучи(1837г) в опыте вторичного сокращения. Нерв 2 препарата лягушки набрасывали на мышцу 1, а нерв первого раздражали током: сокращались обе мышцы. Сокращение второй мышцы происходило в результате раздражения этой мышцы током, возникающим при сокращении 1 мышцы. Все клетки организма имеют заряд – ПП, обеспечиваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различие концентрации является следствием работы ионных насосов и неодинаковой проницаемости клеточной мембраны для разных ионов. При действии раздражителя на клетку возбудимой ткани вначале повышается проницаемость мембраны для натрия и быстро возвращается в норму. Затем то же самое происходит с калием, вследствие чего Na быстро перемещается в клетку, а К+ выходит из клетки согласно электрохимическому градиенту. Возникает процесс возбуждения –ПД. ПД- быстрое колебание величины мембранного потенциала, вследствие активации и инактивации ионных каналов и диффузии ионов в клетку и из клетки. Величина ПД колеблется в пределах 80-130мВ(у нервного 110мВ, у мышечного до 130мВ)Амплитуда ПД не зависит от силы раздражения. Она всегда максимальна для данной клетки в конкретных условиях(Закон все или ничего) Фаза деполяризации: уменьшение заряда клетки до нуля.Она развивается при действии деполяризующего раздражителя на клетку(эл.ток). Открываются ворота натриевых каналов. Когда деполяризация достигает КУД – открывается большое число натриевых каналов и натрий лавинообразно входит в клетку.Фаза инверсии:- изменение заряда клетки на противоположный. Имеет 2 части: восходящую и нисходящую. Восходящая обеспечивается в основном входом натрия в клетку. Нисходящая- закрытие натриевых каналов и выход калия на мембрану. Фаза реполяризации: восстановление ПП. Калий продолжает выходить из клетки по концентрационному градиенту. Теперь клетка снова имеет внутри отрицательный заряд, а снаружи положительный и электрический градиент препятствует выходу калия из клетки. Т.о. вся нисходящая часть ПД обусловлена выходом К+ из клетки.
Теоpия основывается на:
1. Особенностях стpоения биологической мембpаны
2. Устойчивой тpансмембpанной ионной ассиметpии (неодинаковой концентpацией ионов Na + ,K + ,Cl — ,Ca 2+ ,HCO3 — )
Ионную ассиметpию опpеделяют следующие механизмы:
1. Избиpательная пpоницаемость мембpаны для pазличных ионов
2. Работа тpансмембpанных насосов
3. Hаличие силы электpостатического взаимодействия
Катионы К + по концентpационному гpадиенту пассивно диффундиpуют чеpез мембpану из клетки и несут с собой положительный заpяд.
Анионы (глутамат, аспаpтат, сульфаты, оpганические фосфаты, белки и дp.) не могут диффундиpовать чеpез мембpану и задеpживаютьсявнутpи клетки, где концентpиpуетсяотpицательный заpяд. Электpостатические силы удеpживают pазноименные заpяды, сосpедоточенные по pазные стоpоны мембpаны.
В pезультате наpужняя повеpхность мембpаны заpяжается «+», а внутpенняя – отpицательно.
Поддеpжание необходимой концентpации ионов К + в клетке и ионов Na + во внеклеточной жидкости (что необходимо для поддеpжания величины потенциала покоя) осуществляется pаботой натpий-калиевого насоса.
Это обеспечивается пеpеносчиком АТФ-азой с затpатой энеpгии АТФ.
Каждый миофибрилла сост-т из миофиламентов, кот пред-ют собой тонкие нити белка актина и толстые теит белка мибрена. Морф. функцион. единицей мышечных волокон явл. саркомер.
Физиологические свойства скелетных мышц: 1) возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала); 2) низкая проводимость, порядка 10–13 м/с; 3) рефрактерность (занимает по времени больший отрезок, чем у нервного волокна); 4) лабильность; 5) сократимость (способность укорачиваться или развивать напряжение).
Различают два вида сокращения: а) изотоническое сокращение (изменяется длина, тонус не меняется); б) изометрическое сокращение (изменяется тонус без изменения длины волокна). Различают одиночные и титанические сокращения. Одиночные сокращения возникают при действии одиночного раздражения, а титанические возникают в ответ на серию нервных импульсов; 6) эластичность (способность развивать напряжение при растягивании).
ПОТЕНЦИАЛ ДЕЙСТВИЯ




![]()
![]()
Потенциал действия (ПД) — это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала вследствие перемещения ионов в клетку и из клетки и способный распространяться без декремента (без затухания). ПД обеспечивает передачу сигналов между нервными клетками, нервными центрами и рабочими органами; в мышцах ПД обеспечивает процесс электромеханического сопряжения.
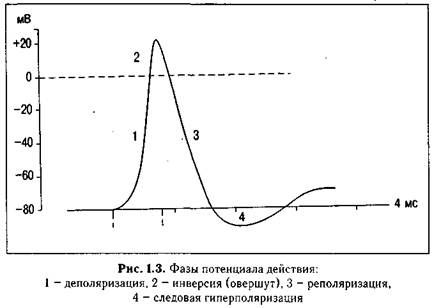
В составе ПД различают четыре фазы:
Естественно, что скорость входа Ыа + в клетку при этом невелика. В этот период, как и во время всей первой фазы (деполяризации), движущей силой, обеспечивающей вход Гч!а + в клетку, являются концентрационный и электрический градиенты. Напомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация № + вне клетки в 10-12 раз больше, чем внутри клетки.
Условием, обеспечивающим вход № + в клетку, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма Ыа-каналов (в некоторых клетках, например, в кардиомиоцитах, в волокнах гладкой мышцы, важную роль в возникновении ПД играют и управляемые каналы для Са 2+ ).

Теперь электрический градиент препятствует входу Ыа + внутрь клетки (положительные заряды отталкиваются друг от друга), Ыа-проводимость снижается. Тем не менее, некоторый период времени (доли миллисекунды) № + продолжает входить в клетку — об этом свидетельствует продолжающееся нарастание ПД. Это означает, что концентрационный градиент, обеспечивающий движение № + в клетку, сильнее электрического, препятствующего входу Ыа + в клетку.
Амплитуда ПД складывается из величины ПП (мембранного потенциала покоящейся клетки) и величины фазы инверсии, составляющей у разных клеток 10-50 мВ. Если мембранный потенциал покоящейся клетки мал, амплитуда ПД этой клетки небольшая.
Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее электрического градиента. Вся нисходящая часть пика ПД обусловлена выходом К + из клетки. Нередко в конце ПД наблюдается замедление реполяризации, что объясняется уменьшением проницаемости клеточной мембраны для К + и замедлением выхода его из клетки из-за частичного закрытия ворот К-каналов. Вторая причина замедления тока К + из клетки связана с возрастанием положительного потенциала наружной поверхности клетки и формированием противоположно направленного электрического градиента.
 |
Роль Са 2+ в возникновении ПД в нервных и мышечных клетках скелетной мускулатуры незначительна. Однако Са 2+ играет важную роль в возникновении ПД сердечной и гладкой мышц, в передаче импульсов от одного нейрона к другому, от нервного волокна к мышечному, в обеспечении мышечного сокращения.
Постепенно проницаемость клеточной мембраны возвращается к исходной (натриевые и калиевые ворота возвращаются в исходное состояние), а мембранный потенциал становится таким же, каким он был до возбуждения клетки. Ыа/К-помпа непосредственно за фазы потенциала действия не отвечает, хотя она и продолжает работать во время развития ПД.
Следовая деполяризация также характерна для нейронов, она может быть зарегистрирована и в клетках скелетной мышцы. Механизм ее изучен недостаточно. Возможно, это связано с кратковременным повышением проницаемости клеточной мембраны для Ыа + и входом его в клетку согласно концентрационному и электрическому градиентам.
В. Запас ионов в клетке, обеспечивающих возникновение возбуждения (ПД), огромен. Концентрационные градиенты ионов в результате одного цикла возбуждения практически не изменяются. Клетка может возбуждаться до 510 5 раз без подзарядки, то есть без работы Ыа/К-насоса.
БИОЛОГИЧЕСКИЙ ОТДЕЛ ЦЕНТРА ПЕДАГОГИЧЕСКОГО МАСТЕРСТВА

Потенциал покоя и потенциал действия
Автор статьи Зыбина А.М.
Потенциал покоя, как и любой мембранный потенциал формируется за счет избирательной проницаемости клеточной мембраны. Как известно, плазмолемма состоит из липидного бислоя, через который движение заряженных молекул затруднено. Белки, встроенные в мембрану, могут избирательно изменять проницаемость мембраны для различных ионов, в зависимости от приходящих стимулов. При этом, для формирования потенциала покоя ведущую роль играют ионы калия, кроме них важны ионы натрия и хлора.
Рис. 1. Концентрации и распределение ионов с внутренней и внешней стороны мембраны.
Большинство ионов распределяются неравномерно с внутренней и внешней стороны клетки (рис. 1). Внутри клетки концентрация ионов калия выше, а натрия и хлора – ниже, чем снаружи. В состоянии покоя мембрана проницаема для ионов калия и практически непроницаема для ионов натрия и хлора. Несмотря на то, что калий может свободно выходить из клетки, его концентрации остаются неизменными благодаря отрицательному заряду на внутренней стороне мембраны. Таким образом, на калий действуют две силы, находящиеся в равновесии: осмотические (градиент концентрации К + ) и электрические (заряд мембраны), благодаря чему число входящих в клетку ионов калия равно выходящим. Движение калия осуществляется через калиевые каналы утечки, открытые в состоянии покоя. Величину заряда мембраны, при которой ионы калия находятся в равновесии можно вычислить по уравнению Нернста:
Чтобы учесть проникновение других ионов в клетку существует уравнение Нернста-Гольдмана:
Потенциал действия (ПД) может возникать в возбудимых клетках. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то ПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет кратковременная перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной, после чего восстановится ПП. Это кратковременное изменение ПП, происходящее при возбуждении клетки называется потенциалом действия.
Рис. 2. Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
На записи ПД выглядит как кратковременный пик (рис. 44), имеющий несколько фаз.
Во время ПД происходит изменение полярности заряда мембраны. Фаза ПД, при которой заряд мембраны положителен, называется овершутом (рис. 2).
Для генерации ПД оказывается очень важной система активации и инактивации потенциал-управляемых натриевых каналов (рис. 3). Эти каналы имеют две створки: активационную (М-ворота) и инактивационную (Н-ворота). В состоянии покоя М-ворота открыты, а Н-ворота закрыты. Во время деполяризации мембраны М-ворота быстро открываются, а Н-ворота начинают закрываться. Ток натрия в клетку возможен пока М-ворота уже открыты, а Н-ворота еще не закрылись. Вход натрия приводит к дальнейшей деполяризации клетки, приводя к открытию большего количества каналов и запуская цепочку положительной обратной связи. Деполяризация мембраны будет продолжаться до тех пор, пока все потенциал-управляемые натриевые каналы не окажутся инактивированными, что происходит на пике ПД. Минимальная величина стимула, приводящая к возникновению ПД называется пороговой. Таким образом, возникший ПД будет подчиняться закону «все или ничего» и его величина не будет зависеть от величины стимула, вызвавшего ПД.
Благодаря Н-воротам инактивация канала происходит раньше, чем потенциал на мембране достигнет равновесной величины по натрию. После прекращения поступления натрия в клетку, происходит реполяризация за счет выходящих из клетки ионов калия. При этом к каналам утечки в этом случае подключаются еще и потениал-активируемые калиевые каналы. Во время реполяризации, в быстром натриевом канале быстро закрываются М-ворота. Н-ворота открываются гораздо медленнее и остаются закрытыми еще некоторое время после возвращения заряда к потенциалу покоя. Этот период принято называть периодом рефрактерности.
Рис. 3. Работа потенциал-управляемого натриевого канала.
Концентрации ионов внутри клетки восстанавливает натрий-калиевая АТФаза, которая с затратой энергии в виде АТФ откачивает из клетки 3 иона натрия и закачивает 2 иона калия.
По немиелинизированному волокну или по мембране мышцы потенциал действия распространяется непрерывно. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до порогового значения, в результате чего на соседнем участке возникает деполяризация. Главную роль в возникновении потенциала на новом участке мембраны предыдущий участок. При этом на каждом участки сразу после ПД наступает период рефрактерности, за счет которое ПД распространяется однонаправленно. При прочих равных условиях распространение потенциала действия по немиелинизированному аксону происходит тем быстрее, чем больше диаметр волокна. У млекопитающих скорость составляет 1-4 м/с. Поскольку у беспозвоночных животных отсутствует миелин, в гигантских аксонах кальмара скорость ПД может достигать 100 м/c.
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах немиелинизированных волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до порогового значения, что приводит к возникновению в них новых потенциалов действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-й, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Сальтаторное проведение увеличивает скорость проведения ПД 15-20 раз до 120 м/с.
Работа нейронов
Нервная система состоит из нейронов и глиальных клеток. Однако, главную роль в проведении и передаче нервных импульсов играют нейроны. Они получают информацию от множества клеток по дендритам, анализируют ее и передают или не передают на следующий нейрон.
Передача нервного импульса с одной клетки на другую осуществляется с помощью синапсов. Различают два основных типа синапсов: электрические и химические (рис. 4). Задача любого синапса – передать информацию с пресинаптической мембраны (мембрана аксона) на постсинаптическую (мембрана дендрита, другого аксона, мышцы или другого органа-мишени). Большинство синапсов нервной системы образуется между окончанием аксонов и дендритами, которые в области синапса образуют дендритные шипики.
Преимущество электрического синапса состоит в том, что сигнал с одной клетки на другую переходит без задержки. Кроме того, такие синапсы не утомляются. Для этого пре- и постсинаптические мембраны соединены поперечными мостиками, через которые ионы из одной клетки могут перемещаться в другую. Однако, существенным минусом такой системы является отсутствие однонаправленной передачи ПД. То есть, он может передаваться как с пресинаптической мембраны на постсинаптическую, так и наоборот. Поэтому, такая конструкция встречается достаточно редко и в основном – в нервной системе беспозвоночных.
Рис. 4. Схема строения химического и электрического синапсов.
Химический синапс весьма распространен в природе. О устроен сложнее, так как необходима система преобразования электрического импульса в химический сигнал, затем, вновь в электрический импульс. Все это приводит к возникновению синаптической задержки, которая может составить 0,2-0,4 мс. Кроме того, может произойти истощение запасов химического вещества, что приведет к утомлению синапса. Однако, такой синапс обеспечивает однонаправленность передачи ПД, что является его главным преимуществом.
Рис. 5. Схема работы (а) и электронная микрофотография (б) химического синапса.
В состоянии покоя окончание аксона, или пресинаптическое окончание, содержит мембранные пузырьки (везикулы) с нейромедиатором. Поверхность везикул заряжена отрицательно, чтобы предотвратить связывание с мембраной, и покрыта специальными белками, и принимающими участие в высвобождении везикул. В каждом пузырьке находится одинаковое количество химического вещества, которое называется квантом нейромедиатора. Нейромедиаторы весьма разнообразны по химическому строению, однако, большинство из них производятся прямо в окончании. Поэтому, в нем могут находиться системы, для синтеза химического посредника, а также аппарат Гольджи и митохондрии.
Постсинаптическая мембрана содержит рецепторы к нейромедиатору. Рецепторы могут быть в виде как ионных каналов, открывающихся при контакте со своим лигандом (ионотропные), так и мембранными белками, запускающими внутриклеточный каскад реакций (метаботропные). Один нейромедиатор может иметь несколько как ионотропных, так и метаботропных рецепторов. При этом, часть из них может быть возбуждающими, а часть – тормозными. Таким образом, реакцию клетки на нейромедиатор будет определять тип рецептора на ее мембране, и разные клетки могут совершенно по-разному реагировать на одно и то же химическое вещество.
Между пре- и постсинаптической мембраной располагается синаптическая щель, шириной 10-15 нм.
При приходе ПД на пресинаптическое окончание, на нем открываются потенциал-активируемые кальциевые каналы и ионы кальция входят в клетку. Кальций связывается с белками на поверхности везикул, что приводит к их транспортировке к пресинаптической мембране с последующим слиянием мембран. После такого взаимодействия нейромедиатор оказывается в синаптической щели (рис. 5) и может связаться со своим рецептором.
Ионотропные рецепторы – это лиганд-активируемые ионные каналы. Это значит, что канал открывается только в присутствии определенного химического вещества. Для разных нейромедиаторов это могут быть натриевые, кальциевые или хлорные каналы. Ток натрия и кальция вызывает деполяризацию мембраны, поэтому такие рецепторы называют возбуждающими. Хлорный ток приводит к гиперполяризации, что затрудняет генерацию ПД. Следовательно, такие рецепторы называют тормозными.
Метаботропные рецепторы к нейромедиаторам относят к классу рецепторов, ассоцированных с G-белками (GPCR). Эти белки запускают разнообразные внутриклеточные каскады реакций, приводящих в конечном итоге либо к дальнейшей передачи возбуждения, либо к торможению.
После передачи сигнала необходимо быстро удалить нейромедиатор из синаптической щели. Для этого в щели присутствуют либо ферменты расщепляющие, нейромедиатор, либо на пресинаптическом окончании или соседних глиальных клетках могут располагаться транспортеры, закачивающие медиатор в клетки. В последнем случае он может использоваться повторно.
Каждый нейрон получает импульсы от 100 до 100 000 синапсов. Одиночная деполяризация на одном дендрите не приведет к дальнейшей передаче сигнала. На нейрон могут приходит одновременно множество как возбуждающих, так и тормозных стимулов. Все они суммируются на соме нейрона. Такая суммация называется пространственной. Далее, может возникнуть или не возникнуть (в зависимости от пришедших сигналов) ПД в области аксонного холмика. Аксонный холмик – это область аксона, примыкающая к соме и обладающая минимальным порогом ПД. Далее импульс распространяется по аксону, конец которого может сильно ветвиться и образовывать синапсы со множеством клеток. Помимо пространственной, существует временная суммация. Она происходит в случае, поступления часто повторяющихся импульсов от одного дендрита.
Помимо классических синапсов между аксонами и дендритами или их шипиками, существуют также синапсы, модулирующие передачу в других синапсах (рис. 6). К ним относят аксо-аксональные синапсы. Такие синапсы способны усиливать или тормозить синаптическую передачу. То есть, если на окончание аксона, образующего аксо-шипиковый синапс, пришел ПД, а в это время по аксо-аксональному синапсу на него пришел тормозный сигнал, высвобождения нейромедиатора в аксо-шипиковом синапсе не произойдет. Аксо-дендритные синапсы могут изменять проведение мембраной ПД на пути от шипика к соме клетки. Также существуют аксо-соматические синапсы, которые могут влиять на суммацию сигнала в области сомы нейрона.
Таким образом, существует огромное многообразие различных синапсов, отличающихся по составу нейромедиаторов, рецепторов и их местоположению. Все это обеспечивает разнообразие реакций и пластичность нервной системы.
Рис. 6. Разнообразие синапсов в нервной системе.